
 Fitness landscapes
Fitness landscapes
At Home in the Universe: The Search for the Laws of Self-Organization and Complexity (Kindle Edition)
Stuart Kauffman (Wikipedia)
- An American medical doctor, theoretical biologist, and complex systems researcher who studies the origin of life on Earth. He was a professor at the University of Chicago, University of Pennsylvania, and University of Calgary. He is currently emeritus professor of biochemistry at the University of Pennsylvania and affiliate faculty at the Institute for Systems Biology. He is best known for arguing that the complexity of biological systems and organisms might result as much from self-organization and far-from-equilibrium dynamics as from Darwinian natural selection as discussed in his book Origins of Order (1993).
Quick takeaway:
- The book’s central thesis is that complexity in general (and life in particular) is an inevitable consequence of self organizing principles that come into play with non-equilibrium systems. He explores the underlying principles in a variety of ways including binary networks, autocatalytic sets, NK models, and fitness landscapes, both static and co-evolving.
- When I was reading this 20 year old book, I had the impression that his work, particularly on how fitness landscapes are explored have direct relevance to the construction of complex systems today. In particular I was struck by how applicable his work with fitness landscapes and NK models would be to the evaluation of the hyperparameter space associated with building Neural Networks.
- Another point that I found particularly compelling is his descriptions of the incalculable size of the high-dimension spaces of combinatorial possibility. The number of potential combinations on even a smallish binary network would take more time in than the universe has to calculate. As such, there need to be mechanisms that allow for a faster, “good enough” evaluation of the space. That’s why we have historical narratives. They describe a path through this space that has worked. As an example, compare tic-tac-toe to chess. In the former, the every possibility in the game space can be known. Chess has too many possibilities, so instead there are openings, gambits, and endgames, discovered by chess masters that come to us as stories.
Notes:
- Chapter 1: At home in the Universe
- In all these cases, the order that emerges depends on robust and typical properties of the systems, not on the details of structure and function. Under a vast range of different conditions, the order can barely help but express itself. (page 19)
- Nonequilibrium ordered systems like the Great Red Spot are sustained by the persistent dissipation of matter and energy, and so were named dissipative structures by the Nobel laureate Ilya Prigogine some decades ago. These systems have received enormous attention. In part, the interest lies in their contrast to equilibrium thermodynamic systems, where equilibrium is associated with collapse to the most probable, least ordered states. In dissipative systems, the flux of matter and energy through the system is a driving force generating order. In part, the interest lies in the awareness that free-living systems are dissipative structures, complex metabolic whirlpools. (page 21).
- The theory of computation is replete with deep theorems. Among the most beautiful are those showing that, in most cases by far, there exists no shorter means to predict what an algorithm will do than to simply execute it, observing the succession of actions and states as they unfold. The algorithm itself is its own shortest description. It is, in the jargon of the field, incompressible. (page 22)
- And yet, even if it is true that evolution is such an incompressible process, it does not follow that we may not find deep and beautiful laws governing that unpredictable flow. For we are not precluded from the possibility that many features of organisms and their evolution are profoundly robust and insensitive to details. (page 23)
- Strikingly, such coevolving systems also behave in an ordered regime, a chaotic regime, and a transition regime. (page 27)
- Note that this reflects our Nomadic (chaotic), Flocking (transition) and Stampeding (ordered) states
- This seemingly haphazard process also shows an ordered regime where poor compromises are found quickly, a chaotic regime where no compromise is ever settled on, and a phase transition where compromises are achieved, but not quickly. The best compromises appear to occur at the phase transition between order and chaos. (page 28)
- Chapter 4: Order for Free
- But evolution requires more than simply the ability to change, to undergo heritable variation. To engage in the Darwinian saga, a living system must first be able to strike an internal compromise between malleability and stability. To survive in a variable environment, it must be stable, to be sure, but not so stable that it remains forever static. Nor can it be so unstable that the slightest internal chemical fluctuation causes the whole teetering structure to collapse. (page 73)
- To survive in a variable environment, it must be stable, to be sure, but not so stable that it remains forever static. Nor can it be so unstable that the slightest internal chemical fluctuation causes the whole teetering structure to collapse. (pg 73)
- It is now well known that in most cells, such molecular feedback can give rise to complex chemical oscillations in time and space. (page 74)
- Olfati-Saber and graph laplacians!
- The point in using idealizations in science is that they help capture the main issues. Later one must show that the issues so captured are not altered by removing the idealizations. (page 75)
- Start with observation, build initial simulation and then measure the difference and modify
- If started in one state, over time the system will flow through some sequence of states. This sequence is called a trajectory (page 77)
- I wonder if this can be portrayed as a map? You have to go through one state to get to the next. In autocatalytic systems there may be multiple systems that may be similar and yet have branch points (plant cells, animal cells, bacteria)
- To answer these questions we need to understand the concept of an attractor. More than one trajectory can flow into the same state cycle. Start a network with any of these different initial patterns and, after churning through a sequence of states, it will settle into the same state cycle, the same pattern of blinking. In the language of dynamical systems, the state cycle is an attractor and the collection of trajectories that flow into it is called the basin of attraction. We can roughly think of an attractor as a lake, and the basin of attraction as the water drainage flowing into that lake. (page 78)
- Also applicable to social and socio-technical systems. The technology changes the connectivity which could change the shape of the landscape
- One feature is simply how many “inputs” control any lightbulb. If each bulb is controlled by only one or two other lightbulbs, if the network is “sparsely connected,” then the system exhibits stunning order. If each bulb is controlled by many other light-bulbs, then the network is chaotic. So “tuning” the connectivity of a network tunes whether one finds order or chaos. The second feature that controls the emergence of order or chaos is simple biases in the control rules themselves. Some control rules, the AND and OR Boolean functions we talked about, tend to create orderly dynamics. Other control rules create chaos. (page 80)
- In our more velocity-oriented system, this is Social Influence Horizon) and is dynamic over time
- Consider networks in which each lightbulb receives input from only one other. In these K = 1 networks, nothing very interesting happens. They quickly fall into very short state cycles, so short that they often consist of but a single state, a single pattern of illumination. Launch such a K = 1 network and it freezes up, saying the same thing over and over for all time. (page 81)
- At the other end of the scale, consider networks in which K = N, meaning that each lightbulb receives an input from all lightbulbs, including itself. One quickly discovers that the length of the networks’ state cycles is the square root of the number of states. Consider the implications. For a network with only 200 binary variables—bulbs that can be on or off—there are 2200 or 1060 possible states. (page 81)
- Such K = N networks do show signs of order, however. The number of attractors in a network, the number of lakes, is only N/e, where e is the basis of the natural logarithms, 2.71828. So a K = N network with 100,000 binary variables would harbor about 37,000 of these attractors. Of course, 37,000 is a big number, but very very much smaller than 2100,000, the size of its state space. (page 82)
- Need to look into if there is some kind of equivalent in the SIH settings
- The order arises, sudden and stunning, in K = 2 networks. For these well-behaved networks, the length of state cycles is not the square root of the number of states, but, roughly, the square root of the number of binary variables. Let’s pause to translate this as clearly as we can. Think of a randomly constructed Boolean network with N = 100,000 lightbulbs, each receiving K = 2 inputs. The “wiring diagram” would look like a madhatterly scrambled jumble, an impenetrable jungle. Each lightbulb has also been assigned at random a Boolean function. The logic is, therefore, a similar mad scramble, haphazardly assembled, mere junk. The system has 2100,000 or 1030,000 states—megaparsecs of possibilities—and what happens? The massive network quickly and meekly settles down and cycles among the square root of 100,000 states, a mere 317. (page 83)
- The reason complex systems exist on, or in the ordered regime near, the edge of chaos is because evolution takes them there. (page 89)
- Chapter 5: The Mystery of Ontology
- Another way to ensure orderly behavior is to construct networks using what are called canalyzing Boolean functions. These Boolean rules have the easy property that at least one of the molecular inputs has one value, which might be 1 or 0, which by itself can completely determine the response of the regulated gene. The OR function is an example of a canalyzing function (Figure 5.3a). An element regulated by this function is active at the next moment if its first, or its second, or both inputs are active at the current moment. Thus if the first input is active, then the regulated element is guaranteed to be active at the next moment, regardless of the activity of the second input. (page 103)
- This is max pooling
- For most perturbations, a genomic system on any attractor will exhibit homeostatic return to the same attractor. The cell types are fundamentally stable. But for some perturbations, the system flows to a different attractor. So differentiation occurs naturally. And the further critical property is this: from any one attractor, it is possible to undergo transitions to only a few neighboring attractors, and from them other perturbations drive the system to still other attractors. Each lake, as it were, is close to only a few other lakes. (page 110)
- Another way to ensure orderly behavior is to construct networks using what are called canalyzing Boolean functions. These Boolean rules have the easy property that at least one of the molecular inputs has one value, which might be 1 or 0, which by itself can completely determine the response of the regulated gene. The OR function is an example of a canalyzing function (Figure 5.3a). An element regulated by this function is active at the next moment if its first, or its second, or both inputs are active at the current moment. Thus if the first input is active, then the regulated element is guaranteed to be active at the next moment, regardless of the activity of the second input. (page 103)
- Chapter 6: Noah’s Vessel
- That we eat our meals rather than fusing with them marks, I believe, a profound fact. The biosphere itself is supracritical. Our cells are just subcritical. Were we to fuse with the salad, the molecular diversity this fusion would engender within our cells would unleash a cataclysmic supracritical explosion. The explosion of molecular novelty would soon be lethal to the unhappy cells harboring the explosion. The fact that we eat is not an accident, one of many conceivable methods evolution might have alighted on to get new molecules into our metabolic webs. Eating and digestion, I suspect, reflect our need to protect ourselves from the supracritical molecular diversity of the biosphere. (page 122)
- We may be discovering a universal in biology, a new law: if our cells are subcritical, then, presumably, so too are all cells—bacteria, bracken, fern, bird, man. Throughout the supracritical explosion of the biosphere, cells since the Paleozoic, cells since the start, cells since 3.45 billion years ago must have remained subcritical. If so, then this subcritical–supracritical boundary must have always set an upper limit on the molecular diversity that can be housed within one cell. A limit exists, then, on the molecular complexity of the cell. (page 126)
- If local ecosystems are metabolically poised at the subcritical–supracritical boundary, while the biosphere as a whole is supracritical? Then what a new tale we tell, of life cooperating to beget ever new kinds of molecules, and a biosphere where local ecosystems are poised at the boundary, but have collectively crept slowly upward in total diversity by the supracritical character of the whole planet. The whole biosphere is broadly collectively autocatalytic, catalyzing its own maintenance and ongoing molecular exploration. (page 130)
- Chapter 8: High-Country Adventures
- what would happen if, in addition to attempting to evolve such a computer program, we were more ambitious and attempted to evolve the shortest possible program that will carry out the task? Such a “shortest program” is one that is maximally compressed; that is, all redundancies have been squeezed out of it. Evolving a serial computer program is either very hard or essentially impossible because it is incredibly fragile. Serial computer programs contain instructions such as “compare two numbers and do such and such depending on which is larger” or “repeat the following action 1,000 times.” The computation performed is extremely sensitive to the order in which actions are carried out, the precise details of the logic, numbers of iterations, and so forth. The result is that almost any random change in a computer program produces “garbage.” Familiar computer programs are precisely the kind of complex systems that do not have the property that small changes in structure yield small changes in behavior. Almost all small changes in structure lead to catastrophic changes in behavior. (page 152)
- This is the inherent problem we are grappling with in our “barely controlled systems”. All the elements involved are brittle and un-evolvable
- It seems likely that there is no way to evolve a maximally compressed program in less time than it would take to exhaustively generate all possible programs, testing each to see if it carries out the desired task. When all redundancy has been squeezed from a program, virtually any change in any symbol would be expected to cause catastrophic variation in the behavior of the algorithm. Thus nearby variants in the program compute very different algorithms. (page 154)
- because the program is maximally compressed, any change will cause catastrophic alterations in the computation performed. The fitness landscape is entirely random. The next fact is this: the landscape has only a few peaks that actually perform the desired algorithm. In fact, it has recently been shown by the mathematician Gregory Chaitin that for most problems there is only one or, at most, a few such minimal programs. It is intuitively clear that if the landscape is random, providing no clues about good directions to search, then at best the search must be a random or systematic search of all the 10300 possible programs to find the needle in the haystack, the possibly unique minimal program. This is just like finding Mont Blanc by searching every square meter of the Alps; the search time is, at best, proportional to the size of the program space. (page 155)
- I’ve been thinking about hyperparameter tuning in the wrong way. There need(?) to be two approaches – one that works in evolvable spaces where there can be gradualism. The other approach cas to work in discontinuous regions, such as what activation function to use.
- The question of what kinds of complex systems can be assembled by an evolutionary search process not only is important for understanding biology, but may be of practical importance in understanding technological and cultural evolution as well. The sensitivity of our most complex artifacts to catastrophic failure from tiny causes—for example, the Challenger disaster, the failed Mars Observer mission, and power-grid failures affecting large regions—suggests that we are now butting our heads against a problem that life has nuzzled for enormously longer periods: how to produce complex systems that do not teeter on the brink of collapse. Perhaps general principles governing search in vast spaces of possibilities cover all these diverse evolutionary processes, and will help us design—or even evolve—more robust systems. (page 157)
- Once we understand the nature of these random landscapes and evolution on them, we will better appreciate what it is about organisms that is different, how their landscapes are nonrandom, and how that nonrandomness is critical to the evolutionary assembly of complex organisms. We will find reasons to believe that it is not natural selection alone that shapes the biosphere. Evolution requires landscapes that are not random. The deepest source of such landscapes may be the kind of principles of self-organization that we seek. (page 165)
- On random landscapes, finding the global peak by searching uphill is totally useless; we have to search the entire space of possibilities. But even for modestly complex genotypes, or programs, that would take longer than the history of the universe. (page 167)
- Things capable of evolving—metabolic webs of molecules, single cells, multicellular organisms, ecosystems, economic systems, people—all live and evolve on landscapes that themselves have a special property: they allow evolution to “work.” These real fitness landscapes, the types that underlie Darwin’s gradualism, are “correlated.” Nearby points tend to have similar heights. The high points are easier to find, for the terrain offers clues about the best directions in which to proceed. (page 169)
- In short, the contribution to overall fitness of the organism of one state of one trait may depend in very complex ways on the states of many other traits. Similar issues arise if we think of a haploid genotype with N genes, each having two alleles. The fitness contribution of one allele of one gene to the whole organism may depend in complex ways on the alleles of other genes. Geneticists call this coupling between genes epistasis or epistatic coupling, meaning that genes at other places on the chromosomes affect the fitness contribution of a gene at a given place. (page 170)
- The NK model captures such networks of epistatic couplings and models the complexity of the coupling effects. It models epistasis itself by assigning to each trait, or gene, epistatic “inputs” from K other traits or genes. Thus the fitness contribution of each gene depends on the gene’s own allele state, plus the allele states of the K other genes that affect that gene. (page 171)
- I find the NK model fascinating because of this essential point: altering the number of epistatic inputs per gene, K, alters the ruggedness and number of peaks on the landscape. Altering K is like twisting a control knob. (page 172)
- This is really important and should also work with graph laplacians. In other words, not only can we model the connectivity, we can model the stiffness
- our model organism, with its network of epistatic interactions among its genes, is caught in a web of conflicting constraints. The higher K is—the more interconnected the genes are—the more conflicting constraints exist, so the landscape becomes ever more rugged with ever more local peaks. (page 173)
- This sounds oddly like how word2vec is calculated. Which implies that all connected neural networks are correlated and epistatic.
- It is these conflicting constraints that make the landscape rugged and multipeaked. Because so many constraints are in conflict, there is a large number of rather modest compromise solutions rather than an obvious superb solution. (page 173)
- Dimension reduction and polarization are a social solution to this problem
- landscapes with moderate degrees of ruggedness share a striking feature: it is the highest peaks that can be scaled from the greatest number of initial positions! This is very encouraging, for it may help explain why evolutionary search does so well on this kind of landscape. On a rugged (but not random) landscape, an adaptive walk is more likely to climb to a high peak than a low one. If an adapting population were to “jump” randomly into such a landscape many times and climb uphill each time to a peak, we would find that there is a relationship between how high the peak is and how often the population climbed to it. If we turned our landscapes upside down and sought instead the lowest valleys, we would find that the deepest valleys drain the widest basins. (page 177)
- The property that the highest peaks are the ones to which the largest fraction of genotypes can climb is not inevitable. The highest peaks could be very narrow but very high pinnacles on a low-lying landscape with modest broad hilltops. If an adapting population were released at a random spot and walked uphill, it would then find itself trapped on the top of a mere local hilltop. The exciting fact we have just discovered is that for an enormous family of rugged landscapes, the NK family, the highest peaks “drain” the largest basins. This may well be a very general property of most rugged landscapes reflecting complex webs of conflicting constraints. (page 177)
- I think this may be a function of how the landscapes are made. The K in NK somewhat dictates the amount of correlation
- Recall another striking feature of random landscapes: with every step one takes uphill, the number of directions leading higher is cut by a constant fraction, one-half, so it becomes ever harder to keep improving. As it turns out, the same property shows up on almost any modestly rugged or very rugged landscape. Figure 8.9 shows the dwindling fraction of fitter neighbors along adaptive walks for different K values (Figure 8.9a) and the increased waiting times to find fitter variants for different K values (Figure 8.9b). Once K is modestly large, about K = 8 or greater, at each step uphill the number of directions uphill falls by a constant fraction, and the waiting time or number of tries to find that way uphill increases by a constant fraction. This means that as one climbs higher and higher, it becomes not just harder, but exponentially harder to find further directions uphill. So if one can make one try per unit time, the rate of improving slows exponentially. (page 178)
- This is very important in understanding how hyperparameter space needs to be explored
- Optimal solutions to one part of the overall design problem conflict with optimal solutions to other parts of the overall design. Then we must find compromise solutions to the joint problem that meet the conflicting constraints of the different subproblems. (page 179)
- Selection, in crafting the kinds of organisms that exist, may also help craft the kinds of landscapes over which they evolve, picking landscapes that are most capable of supporting evolution—not only by mutation alone, but by recombination as well. Evolvability itself is a triumph. To benefit from mutation, recombination, and natural selection, a population must evolve on rugged but “well-correlated” landscapes. In the framework of NK landscapes, the “K knob” must be well tuned. (page 182)
- This is going to be the trick for machine learning
- even if the population is released on a local peak, it may not stay there! Simply put, the rate of mutation is so high that it causes the population to “diffuse” away from the peak faster than the selective differences between less fit and more fit mutants can return the population to the peak. An error catastrophe, first discovered by Nobel laureate Manfred Eigen and theoretical chemist Peter Schuster, has occurred, for the useful genetic information built up in the population is lost as the population diffuses away from the peak. (page 184)
- Eigen and Schuster were the first to emphasize the importance of this error catastrophe, for it implies a limit to the power of natural selection. At a high enough mutation rate, an adapting population cannot assemble useful genetic variants into a working whole; instead, the mutation-induced “diffusion” over the space overcomes selection, pulling the population toward adaptive peaks. (page 184)
- This limitation is even more marked when seen from another vantage point. Eigen and Schuster also emphasized that for a constant mutation rate per gene, the error catastrophe will arise when the number of genes in the genotype increases beyond a critical number. Thus there appears to be a limit on the complexity of a genome that can be assembled by mutation and selection! (page 184)
- We are seeking a new conceptual framework that does not yet exist. Nowhere in science have we an adequate way to state and study the interleaving of self-organization, selection, chance, and design. We have no adequate framework for the place of law in a historical science and the place of history in a lawful science. (page 185)
- This is the research part of the discussion in the iConference paper. Use the themes in the following paragraphs (self organization, selection, etc. ) to build up the areas that need to be discussed and researched.
- The inevitability of historical accident is the third theme. We can have a rational morphology of crystals, because the number of space groups that atoms in a crystal can occupy is rather limited. We can have a periodic table of the elements because the number of stable arrangements of the subatomic constituents is relatively limited. But once at the level of chemistry, the space of possible molecules is vaster than the number of atoms in the universe. Once this is true, it is evident that the actual molecules in the biosphere are a tiny fraction of the space of the possible. Almost certainly, then, the molecules we see are to some extent the results of historical accidents in this history of life. History arises when the space of possibilities is too large by far for the actual to exhaust the possible. (page 186)
- Here is a firm foothold: an evolutionary process, to be successful, requires that the landscapes it searches are more or less correlated. (page 186)
- This is a meta design constraint that needs to be discussed (iConference? Antonio’s workshop?)
- Nonequilibrium systems can be robust as well. A whirlpool dissipative system is robust in the sense that a wide variety of shapes of the container, flow rates, kinds of fluids, and initial conditions of the fluids lead to vortices that may persist for long periods. So small changes in the construction parameters of the system, and initial conditions, lead to small changes in behavior. (page 187)
- Whirlpools are attractors in a dynamical system. Attractors, however, can be both stable and unstable. Instability arises in two senses. First, small changes in the construction of the system may dramatically alter the behavior of the system. Such systems are called structurally unstable. In addition, small changes in initial conditions, the butterfly effect, can sharply change subsequent behavior. Conversely, stable dynamical systems can be stable in both senses. Small changes in construction may typically lead to small changes in behavior. The system is structurally stable. And small changes in initial conditions can lead to small changes in behavior. (page 187)
- We know that there is a clear link between the stability of the dynamical system and the ruggedness of the landscape over which it adapts. Chaotic Boolean networks, and many other classes of chaotic dynamical systems, are structurally unstable. Small changes wreak havoc on their behavior. Such systems adapt on very rugged landscapes. In contrast, Boolean networks in the ordered regime are only slightly modified by mutations to their structure. These networks adapt on relatively smooth fitness landscapes. (page 187)
- We know from the NK landscape models discussed in this chapter that there is a relationship between the richness of conflicting constraints in a system and the ruggedness of the landscape over which it must evolve. We plausibly believe that selection can alter organisms and their components so as to modify the structure of the fitness landscapes over which those organisms evolve. By taking genomic networks from the chaotic to the ordered regime, selection tunes network behavior to be sure. By tuning epistatic coupling of genes, selection also tunes landscape structure from rugged to smooth. Changing the level of conflicting constraints in the construction of an organism from low to high tunes how rugged a landscape such organisms explore. (page 188)
- And so we return to a tantalizing possibility: that self-organization is a prerequisite for evolvability, that it generates the kinds of structures that can benefit from natural selection. It generates structures that can evolve gradually, that are robust, for there is an inevitable relationship among spontaneous order, robustness, redundancy, gradualism, and correlated landscapes. Systems with redundancy have the property that many mutations cause no or only slight modifications in behavior. Redundancy yields gradualism. But another name for redundancy is robustness. Robust properties are ones that are insensitive to many detailed alterations. The robustness of the lipid vesicle, or of the cell type attractors in genomic networks in the ordered regime, is just another version of redundancy. Robustness is precisely what allows such systems to be molded by gradual accumulation of variations. Thus another name for redundancy is structural stability—a folded protein, an assembled virus, a Boolean network in the ordered regime. The stable structures and behaviors are ones that can be molded. (page 188)
- This is why evolution may be the best approach for machine learning hyperparameter tuning
- If this view is roughly correct, then precisely that which is self-organized and robust is what we are likely to see preeminently utilized by selection. (page 188)
- The more rare and improbable the forms that selection seeks, the less typical and robust they are and the stronger will be the pressure of mutations to revert to what is typical and robust. (page 189)
- what would happen if, in addition to attempting to evolve such a computer program, we were more ambitious and attempted to evolve the shortest possible program that will carry out the task? Such a “shortest program” is one that is maximally compressed; that is, all redundancies have been squeezed out of it. Evolving a serial computer program is either very hard or essentially impossible because it is incredibly fragile. Serial computer programs contain instructions such as “compare two numbers and do such and such depending on which is larger” or “repeat the following action 1,000 times.” The computation performed is extremely sensitive to the order in which actions are carried out, the precise details of the logic, numbers of iterations, and so forth. The result is that almost any random change in a computer program produces “garbage.” Familiar computer programs are precisely the kind of complex systems that do not have the property that small changes in structure yield small changes in behavior. Almost all small changes in structure lead to catastrophic changes in behavior. (page 152)
- Chapter 9: Organisms and Artifacts
- Might the same general laws govern major aspects of biological and technological evolution? Both organisms and artifacts confront conflicting design constraints. As shown, it is those constraints that create rugged fitness landscapes. Evolution explores its landscapes without the benefit of intention. We explore the landscapes of technological opportunity with intention, under the selective pressure of market forces. But if the underlying design problems result in similar rugged landscapes of conflicting constraints, it would not be astonishing if the same laws governed both biological and technological evolution. (page 192)
- I begin by describing a simple, idealized kind of adaptive walk—long-jump adaptation—on a correlated but rugged landscape. We have already looked at adaptive walks that proceed by generating and selecting single mutations that lead to fitter variants. Here, an adaptive walk proceeds step-by-step in the space of possibilities, marching steadfastly uphill to a local peak. Suppose instead that we consider simultaneously making a large number of mutations that alter many features at once, so that the organism takes a “long jump” across its fitness landscape. Suppose we are in the Alps and take a single normal step. Typically, the altitude where we land is closely correlated with the altitude from which we started. There are, of course, catastrophic exceptions; cliffs do occur here and there. But suppose we jump 50 kilometers away. The altitude at which we land is essentially uncorrelated with the altitude from which we began, because we have jumped beyond what is called the correlation length of the landscape. (page 192)
- A very simple law governs such long-jump adaptation. The result, exactly mimicking adaptive walks via fitter single-mutant variants on random landscapes is this: every time one finds a fitter long-jump variant, the expected number of tries to find a still better long-jump variant doubles! (page 193)
- Intelligence is computation, and expensive
- As the number of genes increases, long-jump adaptations becomes less and less fruitful; the more complex an organism, the more difficult it is to make and accumulate useful drastic changes through natural selection. (Page 194)
- The germane issue is this: the “universal law” governing long-jump adaptation suggests that adaptation on a correlated landscape should show three time scales—an observation that may bear on the Cambrian explosion. Suppose that we are adapting on a correlated, but rugged NK landscape, and begin evolving at an average fitness value. Since the initial position is of average fitness, half of all nearby variants will be better. But because of the correlation structure or shape of the landscape, those nearby variants are only slightly better. In contrast, consider distant variants. Because the initial point is of average fitness, again half the distant variants are fitter. But because the distant variants are far beyond the correlation length of the landscape, some of them can be very much fitter than the initial point. (By the same token, some distant variants can be very much worse.) Now consider an adaptive process in which some mutant variants change only a few genes, and hence search the nearby vicinity, while other variants mutate many genes, and hence search far away. Suppose that the fittest of the variants will tend to sweep through the population the fastest. Thus early in such an adaptive process, we might expect the distant variants, which are very much fitter than the nearby variants, to dominate the process. If the adapting population can branch in more than one direction, this should give rise to a branching process in which distant variants of the initial genotype, differing in many ways from one another as well, emerge rapidly. Thus early on, dramatically variant forms should arise from the initial stem. Just as in the Cambrian explosion, the species exhibiting the different major body plans, or phyla, are the first to appear. (Page 195)
- Because the fraction of fitter nearby variants dwindles very much more slowly than in the long-jump case. In short, in the mid term of the process, the adaptive branching populations should begin to climb local hills. (Page 195)
- The implication is this: when fitness is average, the fittest variants will be found far away. As fitness improves, the fittest variants will be found closer and closer to the current position. (Page 196)
- So with hyperparameter tuning, change many variables initially, and reduce as the fitness results level out and proceed up the local hill
- Uniting these two features of rugged but correlated landscapes, we should find radiation that initially both is bushy and occurs among dramatically different variants, and then quiets to scant branching among similar variants later on as fitness increases. (page 198)
- Despite the fact that human crafting of artifacts is guided by intent and intelligence, both processes often confront problems of conflicting constraints. (Page 202)
- Dimension reduction is a way of reducing those constraints, but the cost is ignoring the environment. Ideologies must be simple to allow for dense connection without conflict
- As better designs are found, it becomes progressively harder to find further improvements, so variations become progressively more modest. Insofar as this is true, it is obviously reminiscent of the claims for the Cambrian explosion, where the higher taxa filled in from the top down. (Page 202)
- This is a design trap. Since designing for more constraints limits hill climbing, designing for individuals and cultures could make everything grind to a halt. Designing for cultures needs to have a light footprint
- There is something very familiar about this in the context of technological trajectories and learning effects: the rate of finding fitter variants (that is, making better products or producing them more cheaply) slows exponentially, and then ceases when a local optimum is found. This is already almost a restatement of two of the well-known aspects of learning effects. First, the total number of “tries” between finding fitter variants increases exponentially; thus we expect that increasingly long periods will pass with no improvements at all, and then rapid improvements as a fitter variant is suddenly found. Second, adaptive walks that are restricted to search the local neighborhood ultimately terminate on local optima. Further improvement ceases. (Page 204)
- it seems worthwhile to consider seriously the possibility that the patterns of branching radiation in biological and technological evolution are governed by similar general laws. Not so surprising, this, for all these forms of adaptive evolution are exploring vast spaces of possibilities on more or less rugged “fitness” or “cost” landscapes. If the structures of such landscapes are broadly similar, the branching adaptive processes on them should also be similar. (Page 205)
- Chapter 10: An Hour upon the Stage
- The vast puzzle is that the emergent order in communities—in community assembly itself, in coevolution, and in the evolution of coevolution—almost certainly reflects selection acting at the level of the individual organism. (Page 208)
- Models like those of Lotka and Volterra have provided ecologists with simple “laws” that may govern predator-prey relationships. Similar models study the population changes, or population dynamics, when species are linked into more complex communities with tens, hundreds, or thousands of species. Some of these links are “food webs,” which show which species eat which species. But communities are more complex than food webs, for two species may be mutualists, may be competitors, may be host and parasite, or may be coupled by a variety of other linkages. In general, the diverse populations in such model communities might exhibit simple steady-state patterns of behavior, complex oscillations, or chaotic behavior. (Page 211)
- Building an ecology for intelligent machines means doing this. I guess we’ll find out what it’s like to build the garden of eden
- Pimm and his colleagues have struggled to understand these phenomena and have arrived at ideas deeply similar to the models of fitness landscapes we discussed in Chapter 8 and 9. Different communities are imagined as points on a community landscape. Change the initial set of species, and the community will climb to a different peak, a different stable community. (Page 212)
- In these models, Pimm and friends toss randomly chosen species into a “plot” and watch the population trajectories. If any species goes to zero population, hence extinct, it is “removed” from the plot. The results are both fascinating and still poorly understood. What one finds is that, at first, it is easy to add new species, but as more species are introduced, it becomes harder and harder. That is, more randomly chosen species must be tossed into the plot to find one that can survive with the rest of the assembling community. Eventually, the model community is saturated and stable; no further species can be added. (Page 212)
- The community-assembly simulation studies are fascinating for a number of reasons beyond the distribution of extinction events. In particular, it is not obvious why model communities should “saturate,” so that it becomes increasingly difficult and finally impossible to add new species. If one constructs a “community landscape,” in which each point of the terrain represents a different combination of species, then the peaks will represent points of high fitness—combinations that are stable. While a species navigates a fitness landscape by mutating genes, a community navigates a community landscape by adding or deleting a species. Pimm argues that as communities climb higher and higher toward some fitness peak, the ascension becomes harder and harder. As the climb proceeds, there are fewer directions uphill, and hence it is harder to add new species. At a peak, no new species can be added. Saturation is attained. And from one initial point the community can climb to different local peaks, each representing a different stable community. (Page 214)
- In belief spaces, this could help to explain the concept of velocity. It is mechanism for stumbling into new parts of the fitness landscape. And there is something about how ideas go stale.
- In a coevolutionary arms race, when the Red Queen dominates, all species keep changing and changing their genotypes indefinitely in a never-ending race merely to sustain their fitness level. (Page 216)
- This should also apply to belief spaces
- Two main behaviors are imagined. The first image is of Red Queen behavior, where all organisms keep changing their genotypes in a persistent “arms race,” and hence the coevolving population never settles down to an unchanging mixture of genotypes. The second main image is of coevolving populations within or between species that reach a stable ratio of genotypes, an evolutionary stable strategy, and then stop altering genotypes. Red Queen behavior is, as we will soon see, a kind of chaotic behavior. ESS behavior, when all species stop changing, is a kind of ordered regime. (Page 221)
- Just as we can use the NK model to show how genes interact with genes or how traits interact with traits within one organism, we can also use it to show how traits interact with traits between organisms in an ecosystem. (Page 225)
- The ecosystem tends to settle into the ordered, evolutionary stable strategies regime if either epistatic connections, K, within each species are high, so that there are lots of peaks to become trapped on, or if couplings between species, C, is low, so landscapes do not deform much at the adaptive moves of the partners. Or an ESS regime might result when a third parameter, S, the number of species each species interacts with, is low, so that moves by one do not deform the landscapes of many others. (Page 226)
- There is also a chaotic Red Queen regime where the species virtually never stop coevolving (Figure 10.4c). This Red Queen regime tends to occur when landscapes have few peaks to get trapped on, thus when K is low; when each landscape is deformed a great deal by adaptive moves of other species, thus when C is high; or when S is high so that each species is directly affected by very many other species. Basically, in this case, each species is chasing peaks that move away faster than the species can chase them. (Page 228)
- At first, it might seem surprising that low K leads to chaotic ecosystems; in the NK Boolean networks, high K led to chaos. The more inter-couplings, the more likely a small change was to propagate throughout and cause the Boolean system to veer off into butterfly behavior. But with coupled landscapes it is the interconnectedness between the species that counts. When intercoupling, C, is high, moves by one species strongly deform the fitness landscapes of its partners. If any trait in the frog is affected by many traits in the fly, and vice versa, then a small change in traits of one species alters the landscape of the other a lot. The system will tend to be chaotic. Conversely, the ecosystem will tend to be in the ordered regime when the couplings between species, C, is sufficiently low. For much the same reason, if we were to keep K and C the same, but change the number of species S any one species directly interacts with, we would find that if the number is low the system will tend to be ordered, while if the number is high the ecosystem will tend to be chaotic. (Page 228)
- There is something about Tajfel’s opposition identity that might lead to Red Queen scenarios. This would also help to explain the differences between left and right wing behaviours. Right wing is driven by “liberal tears” more than the opposition.
- In fact, the results of our simulations suggest that the very highest fitness occurs precisely between ordered and chaotic behavior! (Page 228)
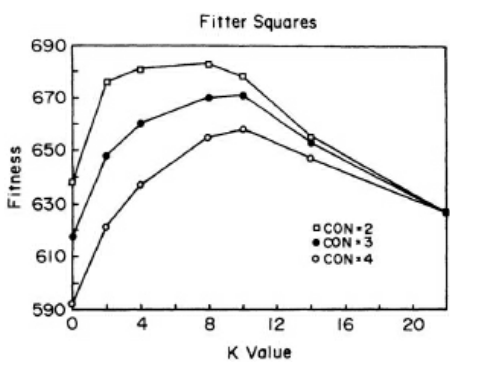
- Tuning an ecosystem. As the richness of epistatic connections between species, K, is increased, tuning the ecosystem from the chaotic to the orderly regime, average fitness at first increases and then decreases. It reaches the highest value midway between the extremes. The experiment is based on a 5 × 5 square lattice ecosystem, in which each of 25 species interacts with at most four other species. (Species on the corners of the lattice interact with two neighbors [CON = 2]; species on the edges interact with three neighbors [CON = 3]; and interior species interact with four neighbors [CON = 4]. N = 24, C = 1, S =25.) (page 229)
- One might start the system with all species having very high K values, coevolving on very rugged landscapes, or all might have very low K values, coevolving on smooth landscapes. If K were not allowed to change, then deep within the high-K ordered regime, species would settle to ESS rapidly; that is, species would climb to poor local peaks and cling to them. In the second, low K, Red Queen chaotic regime, species would never attain fitness peaks. The story no longer stops there, however, for the species can now evolve the ruggedness of their landscapes, and the persistent attempts by species to invade new niches, when successful, will insert a new species into an old niche and may disrupt any ESS attained. (Page 232)
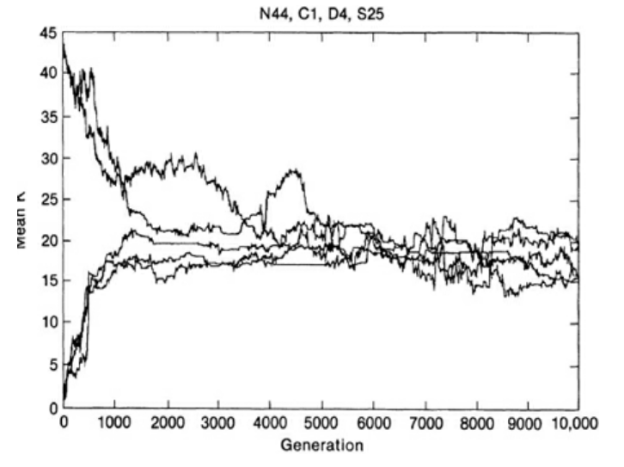
- Figures 10.7 and 10.8 show these results. Each species has N = 44 traits; hence epistatic coupling can be as high as 43, creating random landscapes, or as low as 0, creating Fujiyama landscapes. As generations pass, the average value of K in the coevolving system converges onto an intermediate value of K, 15 to 25, and stays largely within this narrow range of intermediate landscape ruggedness (Above)). Here fitness is high, and the species do reach ESS equilibria where all genotypes stop changing for considerable periods of time, before some invader or invaders disrupt the balance by driving one or more of the coadapted species extinct. (Page 232)
- When K is held high or low, deep in the ordered regime or deep in the chaotic regime, huge extinction avalanches rumble through the model ecosystems. The vast sizes of these events reflect the fact that fitness is low deep in the ordered regime because of the high-K conflicting constraints, and fitness is low deep in the chaotic regime because of the chaotic rising and plunging fitness values. In either case, low fitness of a species makes it very vulnerable to invasion and extinction. The very interesting result is that when the coevolving system can adjust its K range, it self-tunes to values where average fitness is as high as possible; therefore, the species are least vulnerable to invasion and extinction, so extinction avalanches appear to be as rare as possible. This shows up in Figure 10.8, which compares the size distribution and total number of extinction events deep in the ordered regime and after the system has self-tuned to optimize landscape ruggedness, K, and fitness. After the ecosystem self-tunes, the avalanches of extinction events remain a power law—the slope is about the same as when deep in the ordered regime. But over the same total number of generations, far fewer extinction events of each size occur. The self-tuned ecosystem also has far fewer extinction events than does an ecosystem deep in the chaotic regime. In short, the ecosystem self-tunes to minimize the rate of extinction! As if by an invisible hand, all the coevolving species appear to alter the rugged structures of the landscapes over which they evolve such that, on average, all have the highest fitness and survive as long as possible. (Page 234)
- Chapter 11: In Search of Excellence
- Organisms, artifacts, and organizations all evolve and coevolve on rugged, deforming, fitness landscapes. Organisms, artifacts, and organizations, when complex, all face conflicting constraints. So it can be no surprise if attempts to evolve toward good compromise solutions and designs must seek peaks on rugged landscapes. Nor, since the space of possibilities is typically vast, can it be a surprise that even human agents must search more or less blindly. Chess, after all, is a finite game, yet no grand master can sit at the board after two moves and concede defeat because the ultimate checkmate by the opponent 130 moves later can now be seen as inevitable. And chess is simple compared with most of real life. We may have our intentions, but we remain blind watchmakers. We are all, cells and CEOs, rather blindly climbing deforming fitness landscapes. If so, then the problems confronted by an organization—cellular, organismic, business, governmental, or otherwise—living in niches created by other organizations, is preeminently how to evolve on its deforming landscape, to track the moving peaks. (Page 247)
- Evolution is a search procedure on rugged fixed or deforming landscapes. No search procedure can guarantee locating the global peak in an NP-hard problem in less time than that required to search the entire space of possibilities. And that, as we have repeatedly seen, can be hyperastronomical. Real cells, organisms, ecosystems, and, I suspect, real complex artifacts and real organizations never find the global optima of their fixed or deforming landscapes. The real task is to search out the excellent peaks and track them as the landscape deforms. Our “patches” logic appears to be one way complex systems and organizations can accomplish this. (Page 248)
- The basic idea of the patch procedure is simple: take a hard, conflict-laden task in which many parts interact, and divide it into a quilt of nonoverlapping patches. Try to optimize within each patch. As this occurs, the couplings between parts in two patches across patch boundaries will mean that finding a “good” solution in one patch will change the problem to be solved by the parts in the adjacent patches. Since changes in each patch will alter the problems confronted by the neighboring patches, and the adaptive moves by those patches in turn will alter the problem faced by yet other patches, the system is just like our model coevolving ecosystems. Each patch is the analogue of what we called a species in Chapter 10. Each patch climbs toward fitness peaks on its own landscape, but in doing so deforms the fitness landscapes of its partners. As we saw, this process may spin out of control in Red Queen chaotic behavior and never converge on any good overall solution. Here, in this chaotic regime, our system is a crazy quilt of ceaseless changes. Alternatively, in the analogue of the evolutionary stable strategy (ESS) ordered regime, our system might freeze up, getting stuck on poor local peaks. Ecosystems, we saw, attained the highest average fitness if poised between Red Queen chaos and ESS order. We are about to see that if the entire conflict-laden task is broken into the properly chosen patches, the coevolving system lies at a phase transition between order and chaos and rapidly finds very good solutions. Patches, in short, may be a fundamental process we have evolved in our social systems, and perhaps elsewhere, to solve very hard problems. (Page 253)
- It is the very fact that patches coevolve with one another that begins to hint at powerful advantages of patches compared with the Stalinist limit of a single large patch. What if, in the Stalinist limit, the entire lattice settles into a “bad” local minimum, one with high energy rather than an excellent low-energy minimum? The single-patch Stalinist system is stuck forever in the bad minimum. Now let’s think a bit. If we break the lattice up into four 5 × 5 patches just after the Stalinist system hits this bad minimum, what is the chance that this bad minimum is not only a local minimum for the lattice as a whole, but also a local minimum for each of the four 5 × 5 patches individually? You see, in order for the system broken into four patches to “stay” at the same bad minimum, it would have to be the case that the same minimum of the entire lattice happens also to be a minimum for all four of the 5 × 5 patches individually. If not, one or more of the patches will be able to flip a part, and hence begin to move. Once one patch begins to move, the entire lattice is no longer frozen in the bad local minimum. (Page 256)
- Breaking large systems into patches allows the patches literally to coevolve with one another. Each climbs toward its fitness peaks, or energy minima, but its moves deform the fitness landscape or energy landscape of neighboring patches. (Page 257)
- In the chaotic Leftist Italian limit, the average energy achieved by the lattice is only a slight bit less, about 0.47. In short, if the patches are too numerous and too small, the total system is in a disordered, chaotic regime. Parts keep flipping between their states, and the average energy of the lattice is high. (Page 258)
- The answer depends on how rugged the landscape is. Our results suggest that if K is low so the landscape is highly correlated and quite smooth, the best results are found in the Stalinist limit. For simple problems with few conflicting constraints, there are few local minima in which to get trapped. But as the landscape becomes more rugged, reflecting the fact that the underlying number of conflicting constraints is becoming more severe, it appears best to break the total system into a number of patches such that the system is near the phase transition between order and chaos. (Page 258)
- Here, then, is the first main and new result. It is by no means obvious that the lowest total energy of the lattice will be achieved if the lattice is broken into quilt patches, each of which tries to lower its own energy regardless of the effects on surrounding patches. Yet this is true. It can be a very good idea, if a problem is complex and full of conflicting constraints, to break it into patches, and let each patch try to optimize, such that all patches coevolve with one another. (Page 262)
- But what, if anything, characterizes the optimum patch-size distribution? The edge of chaos. Small patches lead to chaos; large patches freeze into poor compromises. When an intermediate optimum patch size exists, it is typically very close to a transition between the ordered and the chaotic regime. (Page 262)
- I’m pretty sure that this can be determined iteratively and within a desired epsilon. It should resemble the way a neural net converges on an accuracy.
- I find it fascinating that hard problems with many linked variables and loads of conflicting constraints can be well solved by breaking the entire problem into nonoverlapping domains. Further, it is fascinating that as the conflicting constraints become worse, patches become ever more helpful. (Page 264)
- I suspect that analogues of patches, systems having various kinds of local autonomy, may be a fundamental mechanism underlying adaptive evolution in ecosystems, economic systems, and cultural systems. (Page 254)
- We are constructing global communication networks, and whipping off into space in fancy tin cans powered by Newton’s third law. The Challenger disaster, brownouts, the Hubble trouble, the hazards of failure in vast linked computer networks—our design marvels press against complexity boundaries we do not understand. (Page 265)
- Patching systems so that they are poised on the edge of chaos may be extremely useful for two quite different reasons: not only do such systems rapidly attain good compromise solutions, but even more essentially, such poised systems should track the moving peaks on a changing landscape very well. The poised, edge-of-chaos systems are “nearly melted.” Suppose that the total landscape changes because external conditions alter. Then the detailed locations of local peaks will shift. A rigid system deep in the ordered regime will tend to cling stubbornly to its peaks. Poised systems should track shifting peaks more fluidly. (Page 266)
- Misspecification arises all the time. Physicists and biologists, trying to figure out how complex biopolymers such as proteins fold their linear sequence of amino acids into compact three-dimensional structures, build models of the landscape guiding such folding and solve for the deep energy minima. Having done so, the scientists find that the real protein does not look like the predicted one. The physicists and biologists have “guessed” the wrong potential surface; they have guessed the wrong landscape and hence have solved the wrong hard problem. They are not fools, for we do not know the right problem. (Page 266)
- Same for Hyperparameter tuning
- We must learn how to learn in the face of persistent misspecification. Suppose we model the production facility, and learn from that model that a particular way to break it into patches is optimal, allowing the system to converge on a suggested solution. If we have misspecified the problem, the detailed solution is probably of little value. But it may often be the case that the optimal way to break the problem into patches is itself very insensitive to misspecifications of the problem. In the NK lattice and patch model we have studied, a slight change in the NK landscape energies will shift the locations of the minima substantially, but may not alter the fact that the lattice should still be broken into 6 × 6 patches. Therefore, rather than taking the suggested solution to the misspecified problem and imposing it on the real facility, it might be far smarter to take the suggested optimal patching of the misspecified problem, impose that on the real production facility, and then try to optimize performance within each of the now well-defined patches. In short, learning how to optimize the misspecified problem may not give us the solution to the real problem, but may teach us how learn about the real problem, how to break it into quilt patches that coevolve to find excellent solutions. (Page 267)
- This is really worth looking at, because it can apply to round tripping simulation and real world systems as well. And a fitness test could be the time to divergence
- receiver-based communication is roughly this: all the agents in a system that is trying to coordinate behavior let other agents know what is happening to them. The receivers of this information use it to decide what they are going to do. The receivers base their decisions on some overall specification of “team” goal. (Page 268)

- This observation suggests that it might be useful if, in our receiver-based communication system, we allowed sites to ignore some of their customers. Let’s say that each site pays attention to itself and a fraction, P, of its customers, and ignores 1 – P of them. What happens if we “tune” P? What happens is shown in Figure 11.8. The lowest energy for the entire lattice occurs when a small fraction of customers is ignored! As Figure 11.8 shows, if each site tries to help itself and all its customers, the system does less well than if each site pays attention, on average, to about 95 percent of its customers. In the actual numerical simulation, we do this by having each site consider each of its customers and pay attention to that customer with a 95 percent probability. In the limit where each site pays attention to no customers, of course, energy of the entire lattice is very high, and hence bad. (Page 268)
- Chapter 12: An Emerging Global Civilization
- Catalytic closure is not mysterious. But it is not a property of any single molecule; it is a property of a system of molecules. It is an emergent property. (Page 275)
- But Fontana found a second type of reproduction. If he “disallowed” general copiers, so they did not arise and take over the soup, he found that he evolved precisely what I might have hoped for: collectively autocatalytic sets of Lisp expressions. That is, he found that his soup evolved to contain a “core metabolism” of Lisp expressions, each of which was formed as the product of the actions of one or more other Lisp expressions in the soup. (Page 278)
- Fontana called copiers “level-0 organizations” and autocatalytic sets “level-1 organizations (Page 279)
- The ever-transforming economy begins to sound like the ever-transforming biosphere, with trilobites dominating for a long, long run on Main Street Earth, replaced by other arthropods, then others again. If the patterns of the Cambrian explosion, filling in the higher taxa from the top down, bespeak the same patterns in early stages of a technological trajectory when many strong variants of an innovation are tried until a few dominant designs are chosen and the others go extinct, might it also be the case that the panorama of species evolution and coevolution, ever transforming, has its mirror in technological coevolution as well? Maybe principles deeper than DNA and gearboxes underlie biological and technological coevolution, principles about the kinds of complex things that can be assembled by a search process, and principles about the autocatalytic creation of niches that invite the innovations, which in turn create yet further niches. It would not be exceedingly strange were such general principles to exist. Organismic evolution and coevolution and technological evolution and coevolution are rather similar processes of niche creation and combinatorial optimization. While the nuts-and-bolts mechanisms underlying biological and technological evolution are obviously different, the tasks and resultant macroscopic features may be deeply similar. (Page 281)
- The difficulty derives from the fact that economists have no obvious way to build a theory that incorporates what they call complementarities. The automobile and gasoline are consumption complementarities. You need both the car and the gas to go anywhere. (Page 282)
- I think that graph laplacians might provide some insight into this behavior
- The use, I claim, is that we can discover the kinds of things that we would expect in the real world if our “as if” mock-up of the true laws lies in the same universality class. Physicists roll out this term, “universality class,” to refer to a class of models all of which exhibit the same robust behavior. So the behavior in question does not depend on the details of the model. Thus a variety of somewhat incorrect models of the real world may still succeed in telling us how the real world works, as long as the real world and the models lie in the same universality class. (Page 283)
- An “enzyme” might be a symbol string in the same pot with a “template matching” (000) site somewhere in it. Here the “enzyme match rule” is that a 0 on the enzyme matches a 1 on the substrate, rather like nucleotide base-pairing. Then given such a rule for “enzymatic sites,” we can allow the symbol strings in the pot to act on one another. One way is to imagine two randomly chosen symbol strings colliding. If either string has an “enzymatic site” that matches a “substrate site” on the other, then the enzymatic site “acts on” the substrate site and carries out the substitution mandated in the corresponding row (Page 285)
- Before we turn to economic models, let us consider some of the kinds of things that can happen in our pot of symbol strings as they act on one another, according to the laws of substitution we might happen to choose. A new world of possibilities lights up and may afford us clues about technological and other forms of evolution. Bear in mind that we can consider our strings as models of molecules, models of goods and services in an economy, perhaps even models of cultural memes such as fashions, roles, and ideas. Bear in mind that grammar models give us, for the first time, kinds of general “mathematical” or formal theories in which to study what sorts of patterns emerge when “entities” can be both the “object” acted on and transformed and the entities that do the acting, creating niches for one another in their unfolding. Grammar models, therefore, help make evident patterns we know about intuitively but cannot talk about very precisely. (Page 287)
- These grammar models also suggest a possible new factor in economic takeoff: diversity probably begets diversity; hence diversity may help beget growth. (Page 292)
-
Diversity begets growth opportunities. Pure growth is fastest in a monoculture of simple items with short maturity cycles
-
 The number of renewable goods with which an economy is endowed is plotted against the number of pairs of symbol strings in the grammar, which captures the hypothetical “laws of substitutability and complementarity.” A curve separates a subcritical regime below the curve and a supracritical regime above the curve. As the diversity of renewable resources or the complexity of the grammar rules increases, the system explodes with a diversity of products. (Page 193)
The number of renewable goods with which an economy is endowed is plotted against the number of pairs of symbol strings in the grammar, which captures the hypothetical “laws of substitutability and complementarity.” A curve separates a subcritical regime below the curve and a supracritical regime above the curve. As the diversity of renewable resources or the complexity of the grammar rules increases, the system explodes with a diversity of products. (Page 193)- Friend, you cannot even predict the motions of three coupled pendula. You have hardly a prayer with three mutually gravitating objects. We let loose pesticides on our crops; the insects become ill and are eaten by birds that sicken and die, allowing the insects to proliferate in increased abundance. The crops are destroyed. So much for control. Bacon, you were brilliant, but the world is more complex than your philosophy. (Page 302)
